Evolution of genetic conflict in plant life cycles
Most plant life cycles involve interactions between tissues belonging to genetically related organisms, such as parents and offspring. These relatives may have different evolutionary interests with regards to resource transfer. Like mammals, many plant groups show extensive post-zygotic provisioning of offspring. When offspring have multiple fathers, there is potential for both parent-offspring and parent-parent conflict over maternal allocation to offspring. Parent-parent conflict can give rise to selection for parent-specific gene expression in offspring (also known as genomic imprinting). Most research in imprinting to date has focused on mammals, but the greater variety of life cycles and breeding systems in plants allow exploration of a broader range of evolutionary questions regarding mechanisms and outcomes of genetic conflict. In collaboration with David Haig, I have been exploring the conflicts that occur among parents and offspring in plant life cycles (doi: 10.1098/rstb.2005.1794 ).
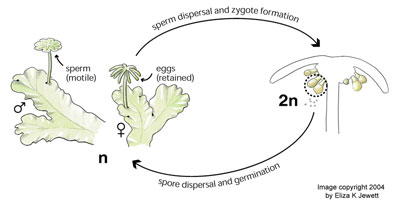 In the life cycles of non-seed-plant bryophytes (mosses, liverworts, hornworts) and pteridophytes (ferns, horsetails, clubmosses), a parasitic diploid plant is nourished by its haploid maternal parent. We have shown that the life cycle exhibited by many bryophytes provides the selective forces necessary for the evolution of genomic imprinting, and that the unusual life cycle of these plants can give rise to a previously undescribed form of sexual conflict. In pteridophytes, on the other hand, the genetic interests of both parents and offspring are aligned, and thus genomic imprinting is less likely to evolve. Our research has provided a potential explanation for some of the striking morphological differences between bryophytes and pteridophytes at the nutritive "placental" junction between the haploid and diploid generations, differences which have been a long-standing mystery in plant morphology (manuscript for submission to Biology Letters, available on request). Future comparative studies could explore how breeding system influences conflict in different bryophyte groups, focusing on the relationship between breeding system and the morphology of the nutritive tissues. In the life cycles of non-seed-plant bryophytes (mosses, liverworts, hornworts) and pteridophytes (ferns, horsetails, clubmosses), a parasitic diploid plant is nourished by its haploid maternal parent. We have shown that the life cycle exhibited by many bryophytes provides the selective forces necessary for the evolution of genomic imprinting, and that the unusual life cycle of these plants can give rise to a previously undescribed form of sexual conflict. In pteridophytes, on the other hand, the genetic interests of both parents and offspring are aligned, and thus genomic imprinting is less likely to evolve. Our research has provided a potential explanation for some of the striking morphological differences between bryophytes and pteridophytes at the nutritive "placental" junction between the haploid and diploid generations, differences which have been a long-standing mystery in plant morphology (manuscript for submission to Biology Letters, available on request). Future comparative studies could explore how breeding system influences conflict in different bryophyte groups, focusing on the relationship between breeding system and the morphology of the nutritive tissues.
Seed plants, including flowering plants, show significant maternal care in the form of seed provisioning. When offspring are sired by more than one father, the kinship theory of genomic imprinting predicts that paternally-expressed (i.e., imprinted) genes should place demands for more resource allocation to individual seeds than maternal plants are selected to give. On the other hand, maternally-expressed genes should be expected to suppress resource transfer from the maternal plant to the seed. The kinship theory further predicts that there will be less conflict between parents over offspring size when levels of polyandry are low and that conflict will disappear under monogamy or selfing. Nonetheless, the importance of breeding system in maintaining imprinting remains an area of great controversy.
 Hermaphroditic plants with closed, obligately inbreeding (cleistogamous) and open, outbreeding (chasmogamous) flowers on the same individual provide a powerful system in which to explore imprinting differences in pollen and ovules that differ only in breeding system, not genetic background. In the closed flowers, the possibility of naturally-occurring polyandry is nil, but this is not the case in the open flowers. The kinship theory then predicts that pollen from open flowers should be more "demanding" than pollen from closed flowers on the same plant; similarly, ovules from closed flowers should be less "resistant" to signals for increased allocation. Such mixed-mating systems have evolved several times in plants, including in the genus Amphicarpaea that I studied for my thesis research. Other genera such as Impatiens and Viola contain suites of species with this floral polymorphism, providing a wealth of systems in which to conduct comparative studies. Controlled, reciprocal crosses among flower types on a single individual as well as among plants can separate changes in seed development and gene expression levels that are attributable to genetic differences or genetic incompatibilities versus those due to epigenetic differences between the flower types. These projects have the potential to verify a strong evolutionary prediction of the kinship theory while at the same time exploring the influence of genetic and epigenetic (e.g. flower type, maternal versus paternal) factors on seed size and seed fitness. Hermaphroditic plants with closed, obligately inbreeding (cleistogamous) and open, outbreeding (chasmogamous) flowers on the same individual provide a powerful system in which to explore imprinting differences in pollen and ovules that differ only in breeding system, not genetic background. In the closed flowers, the possibility of naturally-occurring polyandry is nil, but this is not the case in the open flowers. The kinship theory then predicts that pollen from open flowers should be more "demanding" than pollen from closed flowers on the same plant; similarly, ovules from closed flowers should be less "resistant" to signals for increased allocation. Such mixed-mating systems have evolved several times in plants, including in the genus Amphicarpaea that I studied for my thesis research. Other genera such as Impatiens and Viola contain suites of species with this floral polymorphism, providing a wealth of systems in which to conduct comparative studies. Controlled, reciprocal crosses among flower types on a single individual as well as among plants can separate changes in seed development and gene expression levels that are attributable to genetic differences or genetic incompatibilities versus those due to epigenetic differences between the flower types. These projects have the potential to verify a strong evolutionary prediction of the kinship theory while at the same time exploring the influence of genetic and epigenetic (e.g. flower type, maternal versus paternal) factors on seed size and seed fitness.
Drawing of liverwort life cycle copyright 2004 by Eliza Jewett.
Viola odorata from C.A.M. Lindman's Bilder ur Nordens Flora
(1901-1905)
Home Page |
Site Map |
Contact information
|

